{kind=link}
{kind=link}
{kind=link}

Polymorphisms were identified in mtDNA of Heterobasidion annosum by digesting total genomic DNA with HaeIII, CfoI, or MspI, which recognize the restriction sites GGCC, GCGC, and CCGG, respectively. Most DNA was digested to fragments of less than 2 kb, while AT-rich fragments of 1.7 to 17 kb stood out as bands of uniform intensity after ethidium bromide staining. These fragments hybridized to mtDNA probes and were inherited in a uniparental fashion.
Techniques for identifying polymorphisms in mitochondrial (mt) DNA in fungal populations are generally difficult or expensive. Typically, mtDNA is obtained by separating intact mitochondria from other cell constituents before DNA extraction (Specht et al. 1983 Exp. Mycol. 7:336-343) or by taking advantage of the AT-rich intergenic regions of mtDNA, most commonly by CsCl-bisbenzimide centrifugation (Hudspeth et al. 1980 Biochim. Biophys. Acta 610:221-228). A simpler approach is to use restriction enzymes that selectively digest GC-rich (nuclear) DNA and leave relatively large fragments of multi-copy mtDNA that can be visualized with ethidium bromide staining after electrophoresis (T. D. Bruns and T. E. Chase, unpublished). We developed a procedure that clearly and reproducibly produces discernible AT-rich fragments by digestion of total DNA by HaeIII, CfoI, or MspI (Promega, Madison, Wisconsin), which recognize the restriction sites GGCC, GCGC, and CCGG, respectively. We further showed that such fragments hybridize with mtDNA probes and are inherited in a uniparental fashion indicative of mtDNA. To apply the new technique, total DNA is extracted from cultures grown for 1-2 weeks in liquid medium (DeScenzo and Harrington 1994 Phytopathology 84:534-540). The mycelium is harvested by filtration and finely ground in liquid N2 using a mortar and pestle. Digestions are performed at 37 C in a large volume (300-500 l) of enzyme buffer, with up to 100 ng DNA/ l, 1.0-2.5 mM spermidine, and 5 units of restriction enzyme per microgram of DNA. After 2 h, RNase (A and T1, Sigma Chemical Co.) is added, then the digestion is allowed to proceed for at least 14 h. The digested DNA is concentrated by precipitation (2 volumes of ethanol and 0.2 M NaCl), washed in 70 % ethanol for 2 hr with shaking, centrifuged, the excess ethanol removed with a pipette, and the pellet dried in a Speed Vac. The precipitated DNA is then dissolved in TE (10 mM Tris HCl, 1 mM EDTA) so that the 15 l used for electrophoresis contains 10 g of the original genomic DNA. A large (24 X 19 cm) 1% agarose gel is run at 67 V for 22 h in 1.25X TBE to give good separation of fragments, which are visualized with ethidium bromide staining and UV light.
To test the new technique, 23 isolates of the basidiomycete Heterobasidion annosum were obtained from 22 trees in a small plot in Durham, New Hampshire. This wood decay fungus can saprophytically colonize freshly cut stump tops as basidiospores. Homokaryotic germlings fuse to form dikaryons, which may decay living or dead tree roots and spread to adjacent trees. Somatic incompatibility and DNA fingerprinting with the oligonucleotide (CAT)5 (DeScenzo and Harrington 1994 Phytopathology 84:534-540) identified 14 clones (genets) among the 23 isolates.
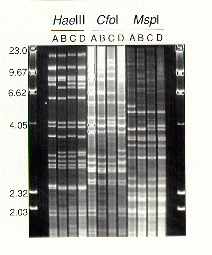
Using each of the three restriction enzymes, four RFLP patterns, designated as mitochondrial haplotypes A, B, C and D (Figure 1), were identified among the 23 H. annosum isolates. All isolates of a clone had identical RFLPs. At least three separate digestions of total DNA were made of each of the isolates, and the same restriction pattern was seen in each digest, although an incompletely digested and faint 5 kb band was found in one of the three replicate runs using HaeIII. Digestions with HaeIII resulted in 28 scorable bands ranging in size from 1.77 to 17 kb. A background smear of stained DNA was evident below 2 kb, but above this, the scored bands stood out clearly and were of uniform intensity. Of the 28 bands, 13 were polymorphic in the population. Digestions with CfoI left some high molecular weight (above 4 kb) background smearing, but 26 bands ranging in size from 1.76 to 14 kb were evident, 11 of which were polymorphic and determined four haplotypes, which corresponded with those identified with HaeIII and MspI digestions (Figure 1). MspI digestions left little background smearing above 2.5 kb, and no bands above 5.3 kb were seen (Figure 1). There were 21 bands greater than 1.7 kb in size that were scored, and eight of these bands were polymorphic.
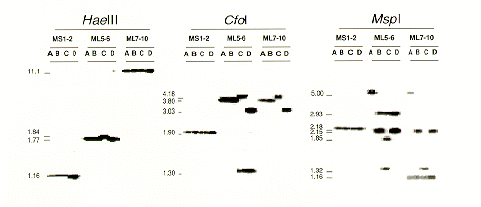
Three specific fragments of the mitochondrial large and small ribosomal subunit genes (L-rDNA and S-rDNA, respectively) were amplified from one of the isolates (B825), labeled and used as hybridization probes to confirm that some of the above fragments were mtDNA. The first and second fragments were amplified from adjacent regions of the L-rDNA using primers ML5/ML6 and ML7/ML10 (ML1D). The third fragment was amplified from the S-rDNA using the primer pair MS1/MS2 (White et al. In Innes et al., eds. 1990 PCR protocols: a guide to methods and applications p. 315-322). Template DNA was obtained without extraction by scraping mycelium from MEA plates (Harrington and Wingfield 1995 Mycologia 87:280-288). Products of 2.2 kb, 1.3 kb and 590 bp, respectively, were purified with the Prep-A-Gene DNA Miniprep Kit (Bio-Rad Laboratories, Hercules, California) and low melting agarose and then radioactivity labeled using 32P dCTP and the Prim-a-Gene (Promega) labeling system. Southern blotting was done according to the method specified (Amersham International, Arlington Heights, Illinois), and hybridization was at 65 C for 16 h. Membranes were washed first with 2X SSC (1X = 150 mM NaCl , 15 mM sodium citrate), 0.1 % SDS for 15 min at 65 C, again for 1 h at 60 C and a third time in 0.2X SSC, 0.1% SDS for 30 min at 60 C. Most of the hybridizing fragments were over 1.7 kb and corresponded to the fragments seen in the ethidium bromide staining (Figure 2). Many of the ML5-6 fragments also hybridized to the ML7-10 probe. Other work (unpublished) has shown that the polymorphisms seen in the L-rDNA is due to a highly variable intron site between the ML5 and ML6 primers.
Further evidence that the ethidium bromide stained bands were mtDNA was found using a multi-locus mtDNA probe, consisting of AT-rich DNA isolated from Armillaria ostoyae using the CsCl-bisbenzimide method and supplied by M. L. Smith and J. B. Anderson (1989 Mycol. Res. 93:247-256). The probe, labeled and used as described above, hybridized with 13 HaeIII fragments, 11 greater than 1.7 kb in size and corresponding to the fragments visualized by ethidium bromide staining. Hybridization signals for bands of 11.1 kb, 4.1 kb, 2.43 kb, 2.39 kb and 1.16 kb were particularly strong and were likely of rDNA, conserved regions expected to show high homology between Armillaria and Heterobasidion. The Armillaria probe hybridized strongly to the 4.18 kb, 3.8 kb, 3.03 kb and 1.9 kb fragments from CfoI digestions, the same fragments that hybridized to the MS1-2, ML5-6 and ML7-10 probes. Likewise, the Armillaria probe hybridized strongly to the MspI fragments of 5.0 kb, 2.93 kb, 2.18 kb, 2.15 kb and 1.32 kb.
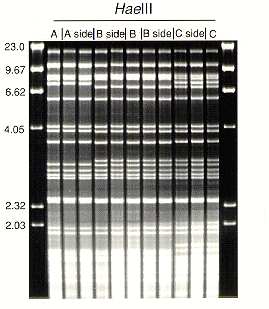
Uniparental inheritance of the ethidium bromide stained fragments was seen in pairings of single-basidiospore strains (homokaryons). Sexually compatible homokaryons of differing mtDNA (haplotypes A, B and C) were paired 10 mm apart on 1 % malt extract agar (MEA) in 90 mm diam plastic Petri dishes for 7 days to form dikaryons. Subcultures were taken from each side of the pairing (from each of the originally homokaryotic mycelia) at about 10-20 mm from the confluence. The subcultures were transferred to MEA plates and inspected for clamp connections. [Secondary mycelia of H. annosum generally grow as mixtures of homokaryotic and dikaryotic (clamped) hyphae.] Individual hyphal tips were taken from the subcultures. Three hyphal tip cultures with clamp connections were analyzed from each side of the pairing plate. The oligonucleotide probe (CAT)5 was used to develop nuclear DNA fingerprints (DeScenzo and Harrington, 1994) of each hyphal tipped culture in order to confirm that the isolates with clamp connections had nuclear markers of both of the homokaryotic parents. In contrast, for each of the three GC restriction enzymes, the banding pattern observed in each dikaryon was identical to that of the respective parent strain from which the dikaryon was subcultured (HaeIII digestions shown in Figure 3). Pairing the homokaryons, therefore, resulted in bi-directional movement of nuclear markers but no movement of mitochondrial markers.
Digestion of total DNA with each enzyme provided numerous and clearly scorable bands in the 1.75 to 15 kb size range that distinguish mtDNA haplotypes. The restriction enzyme HaeIII gave the clearest background and the most scorable bands, although CfoI and MspI digests were also informative. The intensity of the mtDNA fragments was uniform and of much greater intensity than that of the other DNA, which would be expected for multiple copy mtDNA. Hybridizations of the restricted DNA with mtDNA probes indicated that most H. annosum mtDNA is found in fragments above 1.7 kb. The total length of the scorable fragments of the four haplotypes was 46-51 kb with MspI digestions, 91 to 98 kb with CfoI, and 105 to 108 kb with HaeIII, the latter comparable to the mtDNA length estimated for other basidiomycetes, e.g., 80 to 147.6 kb in Armillaria ostoyae (Smith and Anderson 1994 Curr. Genet. 25:545-553). Further support of our contention that the ethidium bromide stained fragments are mtDNA was found in their uniparental inheritance in the synthesis of dikaryons.
Others have used HaeIII and MspI digestions of total fungal DNA to identify AT-rich DNA in an oomycete (Whittaker et al. 1994 Mycol. Res. 98:569-575) and an ascomycete (Freeman et al. 1993 Exp. Mycol. 17:309-322), but those digests did not appear as clear and easy to read as those we present. It is important that digestions be complete in order to remove most of the GC-rich background smear. Relatively pure DNA is needed, and the digestions are carried out overnight with excess restriction enzyme and dilute DNA, which requires precipitation before electrophoresis. The described technique can generate highly reproducible RFLPs representing mt haplotypes in a relatively short time, quicker than any of the other techniques with which we are familiar, and the reagents needed are relatively inexpensive. The technique should prove useful for any fungus that can be grown in culture. In addition to the basidiomycete tested here, we have successfully applied this technique to the ascomycete Ceratocystis virescens and some of its relatives, and a high degree of polymorphism was found.
Figure 1. Ethidium bromide stained agarose gel of HaeIII, CfoI and MspI digestion products of total genomic DNA from four isolates of Heterobasidion annosum representing four mitochondrial haplotypes (A, B, C and D). Sizes of markers in the first and last lanes are given in kb.
Figure 2. HaeIII, CfoI and MspI digestion products of four mitochondrial haplotypes (A, B, C and D) of Heterobasidion annosum hybridized with portions of the small (MS1-2) and large (ML5-6 and ML7-10) subunits of mitochondrial rDNA. Fragment sizes are given in kb.
Figure 3. Ethidium bromide stained agarose gel of HaeIII digestion products of parent homokaryons A, B and C and subcultured dikaryons taken for A X B and B X C pairings. Subcultured dikaryons from the A side of the pairing showed the pattern of the A parent, those from the B side the pattern of the B parent, and those from the C side the pattern of the C parent. Fragment sizes of markers are given in kb.
Return to the FGN43 Table of Contents
Return to the FGSC Home page
Last modified 7/25/96 KMC