{kind=link}

Aramayo, R., and Metzenberg, R. L. - Department of Biomolecular Chemistry. University of Wisconsin. 690 M.S.C., 1300 University Avenue, Madison, WI 53706
We report the construction of pRAUW122, a plasmid capable of directing the integration of gene-constructs by gene replacement at the his-3 locus of Neurospora crassa. Gene targeting at his-3 is selected by growing the transformants in histidine-free medium. The elimination of a second selectable marker (Bmlr), flanking the gene construct, is detected by screening for benomyl sensitivity of the his-3+ transformants. Therefore, his-3+ Bmls transformants have a high probability of having arisen by the desired gene replacement event.
Gene-directed integration is a powerful tool for the construction of strains that are isogenic except for the known differences in the introduced DNA sequences. Integrative transformation at a determined chromosomal position (e.g. the his-3 locus) thus allows the comparison of the level of expression of a specified gene with that of other genes or with different versions of the same gene in the same chromosomal environment. Two loci have routinely been used for gene targeting in Neurospora crassa (am and his-3). Gene replacement at the am locus was developed by Miao et al. (Mol. Cell. Biol. 14: 7059-7067) for the evaluation of potential methylation signals. Gene-directed integration at the his-3 locus was first developed by Karl Hager and Mike Plamann and was based on the plasmid pH303 (K. Hager, Yale University, unpublished observations). Plasmid pH303 and its derivatives integrate at his-3 by a single crossover. This kind of integration generates a functional and a non-functional version of his-3, with the sequences targeted to this locus lying between (viz., his-3+::geneX::his-3). A potential problem with this kind of integration is that the presence of linked direct repeats is accompanied by a non-zero level of vegetative and pre-meiotic recombination between the repeats, with consequent deletion of the sequences between them. In addition, RIP (repeat-induced point mutation), a pre-meiotic process that mutates duplicated DNA sequences, is likely to occur in crosses of such transformants (Selker et al., 1987 Cell 51: 741-752; Selker and Stevens, 1987 Mol. Cell. Biol. 7: 1032-1038; Selker and Garrett, 1988 Proc. Natl. Acad. Sci. USA 85: 6870-6874; Selker, 1990 Annu. Rev. Genet. 24: 579-613; Bowring and Catcheside, 1991 Mol. Gen. Genet. 229: 273-277; Irelan et al., 1994 Genetics 138: 1093-1103). Thus, despite its popular use (Legerton and Yanofsky, 1985 Gene 39: 129-140; Ebbole, 1990 Fungal Genet. Newsl. 37: 15-16; Sachs and Ebbole, 1990 Fungal Genet. Newsl. 37: 35-36; Madi et al., 1994 P.N.A.S. USA 91: 6226-6230; Corrochano et al., 1995 Dev. Biol. 167: 190-200; Luo et al., 1995 Mol. Cell. Biol. 15: 5235-5245) the use of this kind of integration has certain shortcomings.
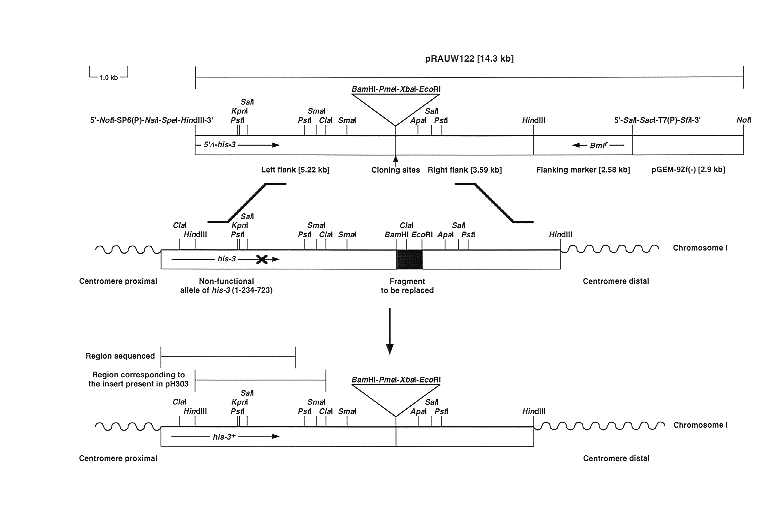
We therefore developed a second generation of plasmids based on a plasmid designated pRAUW122 (Figure 1). Portions of this plasmid and its derivatives can be integrated at his-3 by a double crossover event (e. g., to give his-3+::geneX). The occurrence of the first crossover occurs between the truncated version of his-3 (5' -his-3, HindIII-BamHI fragment - unable to confer histidine independence when integrated ectopically, left flank, Figure 1) and the mutant allele of his-3 (1-234-723) present in the genome and whose mutation was mapped to be somewhere downstream of the SalI restriction site. This event regenerates a functional copy of his-3, which confers histidine prototrophy on the transformant. The second crossover must take place somewhere between the right flank of the construct (EcoRI-HindIII fragment , right flank, Figure 1) and the homologous region in the genome. Its occurrence completes the integration of the his-3 sequences and potential insert and eliminates the downstream portion of the construct, the benomyl resistance gene marker - Bml r, a mutant allele of the Neurospora tub-2 (which is detected as benomyl sensitivity of the transformant), and plasmid sequences (see Figure 1). Consequently, a his-3+, Bml s transformant has a high probability of containing the desired gene replacement, but the event should still be confirmed by molecular analysis. This kind of site-directed gene replacement generates a single functional copy of his-3 without generating linked duplications, thus eliminating the risk of RIP and/or recombination events that could excise the region between repeats. This procedure was based on methods previously developed for mice (Mansour et al., 1988 Nature 336: 348-352; Frohman and Martin, 1989 Cell 56: 145-147) and Neurospora (Aronson et al., 1994 242: 490-494).
Experimental
Plasmid pRAUW122 (Figure 1) was constructed as follows: first, we identified a 9.51 kb HindIII fragment from cosmid C6:4E (Orbach and Sachs, 1991 Fungal Genet. Newsl. 38: 97) that contains the 3'-end of his-3 and a previously unmapped downstream segment of the region. This fragment (unable to confer histidine independence when integrated ectopically) was cloned into the HindIII site of pBC KS(+) (Stratagene, La Jolla, CA) to generate plasmids pRAUW111 and pRAUW112. These plasmids contain inserts in opposite orientations and both were used to generate a basic restriction map of the region. Second, we cloned the 2.58 kb SalI-HindIII fragment, containing the Bmlr gene from pMO63 into the SalI and HindIII sites of pGEM9Zf(-) (Promega, Madison, WI) to generate pRAUW114. Plasmid pMO63 contains a modified version of the Bmlr gene from N. crassa, which lacks the internal BamHI and EcoRI restriction sites and was generously given to us by Marc Orbach (Orbach et al., 1986 Mol. Cell. Biol. 6: 2452-2461). Third, the 9.51 kb HindIII fragment from pRAUW111 was inserted into the HindIII site of pRAUW114 to generate pRAUW116. The Bmlr gene is downstream of the truncated version of his-3. The direction of transcription of his-3 and Bmlr are convergent (see Figure 1). Finally, pRAUW122 was constructed by deleting the 0.7 kb BamHI-EcoRI fragment located approximately in the middle of the 9.51 kb HindIII fragment of pRAUW116 (the ClaI site present within this fragment was consequently lost). The BamHI and EcoRI ends were then ligated in the presence of two oligonucleotides (ORAUW24 [5'-GATCCGTTTAAACTCTAGACCG-3'] and ORAUW25 [5'-AATTCGGTCTAGAGTTTAAACG-3']) whose pairing was designed to regenerate restriction sites for BamHI, PmeI, XbaI, and EcoRI, respectively (see Figure 1). These unique restriction sites, located approximately in the middle of the 9.51 kb HindIII fragment, were included as cloning sites for constructs to be targeted to the his-3 locus. Plasmids pRAUW123 and pRAUW124 both contain a 12.5 kb insert corresponding to the Asm-1 (Ascospore maturation-1) region and will be described in Aramayo et al. (submitted for publication). The final sizes of plasmids pRAUW122, pRAUW123 and pRAUW124 are 14.3 kb, 26.8 kb, and 26.8 kb, respectively.
For production of mycelia and conidia, strains were grown in 1 ml of appropriately supplemented Vogel's Medium N with 2% sucrose, or in 20 ml batches solidified with 1.5% agar in 250 ml Erlenmeyer flasks (Davis and de Serres, 1970 Meth. Enzymol. 17A: 79-143). When supplements were appropriate or required, they were generally used at the following concentrations: L-histidine, 209.6 g/ml; Ca2+/-pantothenate, 10 g/ml; benomyl, 1 g/ml. All platings were done on Vogel's salts with the sugar mixture of Brockman and de Serres to induce colonial growth (Brockman and de Serres, 1963 Am. J. Botany 50: 709-714). Strains were grown for 2 days at 34•C (dark) and 1 day at room temperature (light).
Figure 1. Gene replacements at the his-3 locus of Neurospora crassa. A restriction map of pRAUW122 (linearized with NotI) and a schematic diagram of its integration by gene replacement at the his-3 locus is presented. DNA fragments corresponding to the left and right flanks, flanking marker and pGEM-9Zf(-) sequences of pRAUW122, as well as the position of the restriction sites used to clone our constructs (polylinker) are indicated below the restriction map of the plasmid. 5' -his-3 corresponds to the truncated allele of his-3 that lacks its 5'-end. Bmlr, corresponds to the benomyl resistance allele of the tub-2 gene. The restriction map of the genomic recipient sequences for the gene replacement (the his-3 [1-234-723] allele and downstream region), are indicated below the diagram of pRAUW122. The putative region of the inactivating mutation of the his-3 (1-234-723) allele is indicated by an "x". The hatched area corresponds to the BamHI-EcoRI fragment that will be replaced during the integration. The predicted restriction map generated by the gene replacement event, the approximate location of the sequenced region, and the location of the insert present in pH303 is presented below the downward arrow. Horizontal arrows indicate direction of transcription.
Conidial spheroplasts preparation and transformation were done by a modification of the combined methods of Schweizer et al. (1981 Proc. Natl. Acad. Sci. USA 78: 5086-5090); Akins and Lambowitz (1985 Mol. Cell. Biol. 5: 2272-2278); and Vollmer and Yanofsky (1986 Proc. Natl. Acad. Sci. USA 34: 573-577). Our method (transformation of conidial spheroplasts instead of germlings) represent a significant modification of standard procedures and results in a considerable saving of time. Typically, four 250 ml Erlenmeyer flasks, each containing 20 ml of solid medium, were inoculated and allowed to grow for 3 days at 34 C and 2 days at room temperature. Conidia were harvested into 200 ml of sterile water and filtered through Miracloth. Conidial suspensions were centrifuged at 2,000 x g for 15 min and the pellets were suspended in 20 ml of 1 M aqueous sorbitol. Bovine serum albumin (60 mg) and Novozyme 234 (100 mg) were dissolved in 4 ml of 1 M sorbitol and added to the suspension, which was then shaken at 80 rpm, 30 C, for 90 min. The resulting spheroplasts were then sedimented at 3,500-4,000 x g for 10 min and washed three times with 20 ml of 1 M sorbitol and twice with STC (1 M sorbitol; 50 mM Tris-HCl, pH 8.0; 50 mM CaCl2). The final suspension of spheroplasts was titered with a hemocytometer and adjusted to 2 x 108/ml with STC. For each milliter of spheroplasts, the following volumes of reagents were added: 12.5 ul of DMSO, 62.5 ul of heparin (5 mg/ml in STC) and 250 ul of PTC (40% polyethylene glycol (3355); 50 mM Tris-HCl, pH 8.0; 50 mM CaCl2). The spheroplasts were distributed into samples at 0 C and stored at -70 C.
The fungal strains used in these experiments were RLM 80-21 (mepr his-3; mtr ; Asm-1 ::mtr+; pan-2 A) and RLM 80-23 (mepr his-3; mtr ; Asm-1 ::mtr+; pan-2 a), with the following allele numbers: his-3 (1-234-723), mepr (10), mtr (deletion SR62), and pan-2 (B3[Y153M96]). The construction of the Asm-1 ::mtr+, allele will be described in Aramayo et al. (submitted for publication). We used 5 ug or 20 ug of either linear or circular plasmid DNA (as indicated) and 1.5 x 107 spheroplasts per transformation. Typically, DNA + STC were mixed in a 1.7 ml Eppendorf tube to give a final volume of 250 ul. Spheroplasts (100 ul) were then added to the cold DNA + STC solution. The contents were mixed and incubated on ice for 30 min, after which 1 ml of PTC solution was added to the DNA + STC + spheroplast suspension. Tubes were inverted several times and incubated at room temperature for 30 min. The transformation mixture was then transferred to a 15 ml Falcon tube containing 10 ml of warm selective top-agar (Vogel's sucrose medium + 0.7% agar) and the contents were plated onto one Petri dish. A typical yield was about 3 transformants per microgram. Positive and negative controls consisted of spheroplasts transformed with either 10 g of circular pH303 plasmid or no plasmid DNA, respectively.
Results and Discussion
To determine the frequency of transformation by linear versus circular plasmid DNA, we transformed both hosts, A (RLM 80-21) and a (RLM 80-23) with either linear or circular plasmids pRAUW122, pRAUW123, and pRAUW124. In all cases we obtained a higher transformation frequency using linear than using circular DNA, and the transformation frequencies were independent of the mating type of the host (Table 1).
We selected 36 transformants, 18 obtained with linear pRAUW123 and 18 transformants obtained with linear pRAUW124 (in each case, nine of each mating type) and analyzed them by Southern blotting. Among these, 16 (eight of each mating type), obtained with plasmid pRAUW123, and 17 (nine and eight of mating types A and a, respectively), obtained with plasmid pRAUW124, presented a correct gene replacement event. Among those, eight (four of each mating type), obtained with plasmid pRAUW123, and six (three of each mating type), obtained with plasmid pRAUW124, were isolated in the homokaryotic state without further purification. The other 19 transformants were heterokaryons with varying nuclear ratios. One transformant contained an unpredicted event that corresponded to a site-directed gene replacement, but with plasmid sequences also integrated. Finally, two transformants had a gene replacement that did not include the insert, suggesting that both the first and second crossovers occurred in the left flank of the plasmid. All the transformants described above were tested and found to be sensitive to benomyl, as predicted by Southern data, and confirming the elimination of the flanking marker (Figure 1). Roughly similar results were obtained by transforming with pRAUW122 (data not shown).
Table 1. Transformation results (numbers in parentheses refer to notes at the end)
Plasmid Form First transformation(1) Second transformation(2)
RLM 80-21(3) RLM 80-233(3) RLM 80-21(3) RLM 80-23(3)
pRAUW122 Linear(4) 15.2 +/- 4.3 11.4 +/- 3.4 24.2 +/- 6.2 24.8 +/- 6.4
Circular 2.6 +/- 1.0 2.4 +/- 1.8 3.6 +/- 2.0 4.8 +/- 2.6
pRAUW123 Linear(4) 17.4 +/- 8.3 14.8 +/- 3.3 46.0 +/- 10.6 39.0 +/- 12.8
Circular 0.80 +/- 0.74 0.80 +/- 0.74 2.0 +/- 1.1 2.0 +/- 1.3
pRAUW124 Linear(4) 10.8 +/- 3.2 12.6 +/- 5.5 45.4 +/- 5.3 35.6 +/- 9.0
Circular 0.60 +/- 0.80 0.40 +/- 0.80 1.4 +/- 1.7 2.2 +/- 1.2
(1) Number of transformants/5 ug DNA.
(2) Number of transformants/20 ug DNA. Each number represents the average of 5
independent transformations +/- the standard deviation.
(3) Strain are described in the text
(4) Plasmids were linearized by digesting them with NotI. After digestion,
the enzyme was heat-inactivated and DNA was used for transformation
without further treatment.
We tested 66 transformants obtained with circular DNA for resistance or sensitivity to benomyl. Among them, only four were benomyl resistant, indicating that the predominant event was gene replacement and not gene-directed integration of an entire plasmid.
In summary, pRAUW122 and its derivatives can be used to direct the integration of constructs to the his-3 locus. This is facilitated by the high transforming efficiency of linearized plasmids and by the fact that, under our experimental conditions, transformants homokaryotic for the desired event are obtained at a high frequency.
Acknowledgements
This work was supported by U. S. Public Health Service grant GM08995 to R. L. M.
Return to the FGN43 Table of Contents
Return to the FGSC Home page
Last modified 7/25/96 KMC