A series of six compact fungal transformation vectors containing
polylinkers with multiple unique restriction sites
Martin L. Pall and Joseph P. Brunelli - Department of Genetics and
Cell Biology and Department of Biochemistry/Biophysics, Washington
State University, Pullman, WA 99164-4234
In comparison with transformation vectors available for use in
E. coli or yeast, there has been relatively little development of vectors
for use in filamentous fungi. For example, expression yeast vectors
carrying polylinkers flanked by promoters and terminators are
available for various uses but such vectors have not been in the public
domain for researchers working with filamentous fungi. In addition,
in E. coli, many of the most useful, compact vectors carry polylinkers
into which insertion can be monitored using blue/white screening but
we are only aware of one such vector designed for use in filamentous
fungi (Nelson and Metzenberg, 1992 Fungal Genet. Newsl. 39:59-60).
The most compact available fungal selectable marker is the
Ignite/basta-resistance (bar) gene (Avalos et al. 1989 Curr. Genet.
16:369-372; Straubinger et al. 1992 Fungal Genet. Newsl. 39:82-83;
Pall, this volume). We report here on the construction of six compact
fungal transformation vectors carrying the bar gene. Four of these
carry polylinker regions subcloned from pGEM5Zf(+), pGEM7Zf(+),
and pBluescriptII KS+ (Promega Biotech and Stratagene). The other
two are expression vectors with numerous useful features.
The first four of the vectors are shown in Figure 1 and
described in the text. Three of them (pBARGEM5-1, pBARGEM7-1,
and pBARKS1) are each about 4.5 kb and all four carry the bar gene
under control of the Aspergillus nidulans trpC promoter. The
polylinkers from the pGEM and Bluescript vectors are shown at the
top of each restriction map, with the four to eight sites that are unique
in each construct underlined.
Two important points about these four vectors are as follows:
Insertion of markers into these polylinkers will, in most cases, disrupt
the function of the lacZ` gene which can be monitored by blue/white
screening in medium containing Xgal using such E. coli host strains as
DH5` or SURE. These vectors all replicate at relatively high copy
number in E. coli, allowing relatively easy isolation of their DNA.
One vector, pBARGEM7-2, shown in (Figure 1) carries the lox NotI
lox sequence derived from the pSE936 vector of Elledge et al. (1991,
Proc. Natl. Acad. Sci. USA 88:1731-1735), as shown in the the left top
part of its restriction map. This sequence has several important
features that are described below.
Two expression vectors were constructed carrying the N. crassa mtr or,
alternatively, the A. nidulans gpdA promoter and the A. nidulans trpC
terminator flanking a polylinker with multiple unique sites. The
BamHI to KpnI region of the polylinkers was subcloned from
pBluescriptII KS+. The mtr promoter cloned into pBARMTE1 (fig.
2) is composed of nucleotides 1 through 306 (SalI site) of the insert of
the pCVN2.9 plasmid described previously by Koo and Stuart (1991,
Genome 30:198-203). It was prevously thought that the large open
reading frame of the mtr gene started at position 975 (Koo and Stuart,
op cit) of this sequence but Davin Dillon and David Stadler (personal
communication) have recently shown that it actually starts upstream,
possibly at position 347. The sequence of the gpdA promoter cloned
into pBARGPE1 was from position -679 to position +53 (Punt et al.
1988 Gene 69:49-57). Both expression vectors are designed to express
genes or cDNAs under their own initiation codons. In order to test
the activity of these promoter/terminator combinations, precursor
plasmids of pBARMTE1 and pBARGPE1, containing the same
promoter/terminator sequences but without any fungal selectable
markers were taken and the coding sequence of the bar gene devoid
of any other promoter or terminator activity was inserted into their
polylinkers. These were then used to transform N. crassa, selecting for
Ignite/basta-resistance. Both of these constructs were active in
generating basta-resistant transformants although about 2 to 3 times
as many transformants were produced by the mtr construct as by the
gpdA construct. We infer that both of these promoter/terminator
combinations are active in N. crassa but that the mtr construct may be
somewhat more active than is the gpdA construct.
The lox-NotI-lox sequence, found above in pBARGEM7-2, is present
in the upper left region of the pBARMTE1 and pBARGPE1 plasmid
maps (Figure 2). This sequence has been used by Elledge et al. (1991
Proc. Natl. Acad. Sci. USA 88:1731-1735) and by us (Brunelli and Pall,
unpublished) to attach plasmids to the NotI containing arms of a
derivative of lambda-gt7. This converts a plasmid vector into a lambda
vector of a special type. Because the lox sites are sites of site-specific
recombination catalyzed by the Cre protein, lambda infection of E. coli
strains carrying a cre gene leads to excision of the plasmid sequence
from the lambda vector. This "automatic excision" of the plasmid gives
these vectors the advantages of both lambda and plasmids - the
efficient packaging, infection and storage of large inserts in lambda -
and the compact size and easy handling of plasmids. The EcoRI and
XhoI sites in the polylinker of these vectors are unique not only in the
plasmid versions but also in the lambda versions, allowing these to be
used for construction of genomic and cDNA libaries. We expect to be
inserting each of these three lox-NotI-lox-containing plasmids into
lambda shortly.
The plasmids described are available through the Fungal Genetics
Stock Center.
The bar gene in these constructs is active in E. coli, producing
resistance to 400 micrograms/ml of Ignite/basta added to mineral
salts/glucose media.
We plan to replace the bar gene in some these constructs with the
hygromycin B resistance gene (hph or hyg) to provide an alternative
selectable marker and also change the pattern of unique sites in the
polylinkers.
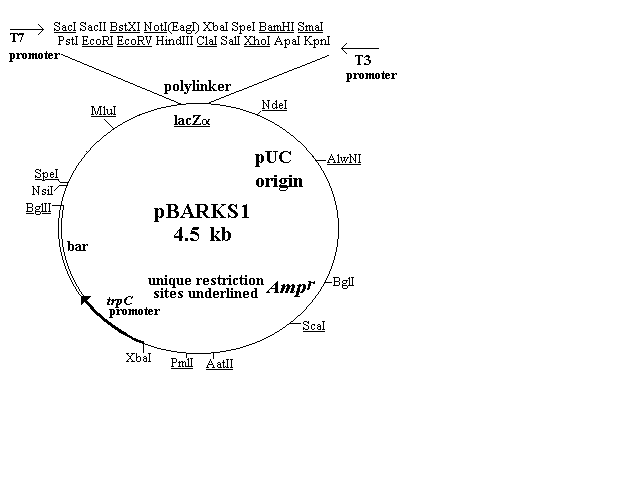
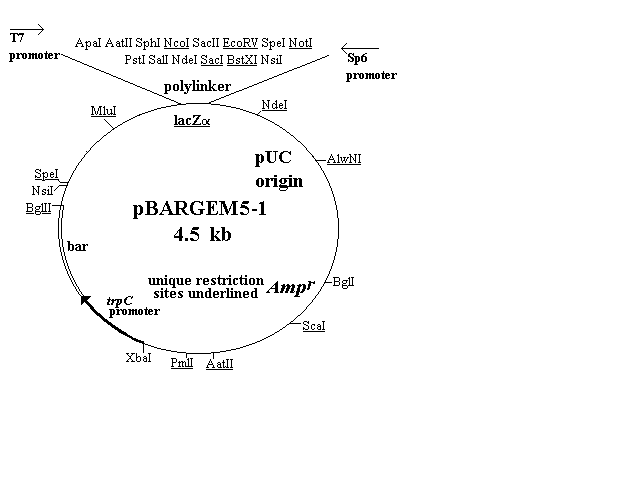

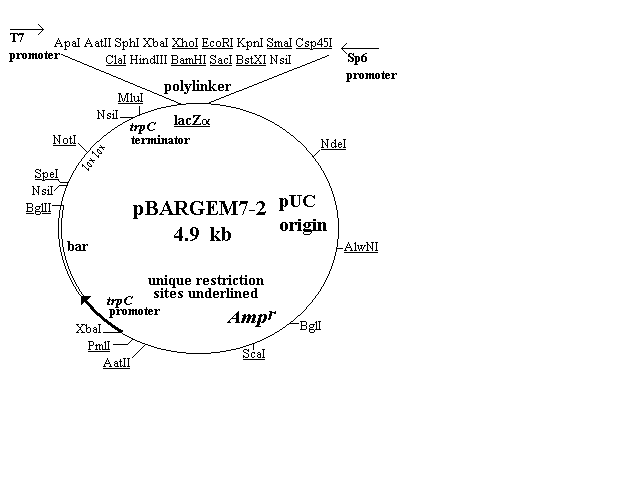
Figure 1. Plasmid maps of four fungal transformation vectors. Each
carries the bar gene as a selectable marker (lower left portion of map),
a polylinker and lacZalpha sequence from pGEM or pBluescript vectors,
including promoters from T3, T7 and/or SP6, and ampicillin-resistance
gene and pUC type origin for growth and selection in E. coli. The
pBARGEM7-2 plasmid also contains the lox-NotI-lox sequence
allowing for attachment to lambda and site-specific recombination
between lox sites.
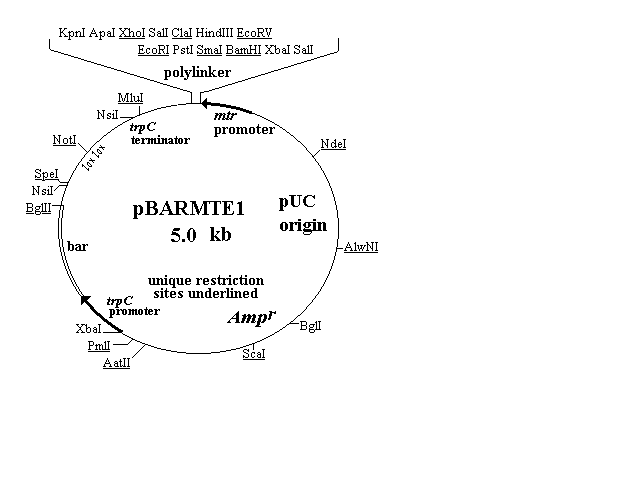
 Figure 2. Plasmid maps of two fungal expression vectors. Each
plasmid carries a polylinker with 5 or 6 unique restriction sites flanked
on the right by a fungal promoter directing transcription into the
polylinker and flanked on the left by the trpC terminator. Both
plasmids carry a bar selectable marker and a lox-NotI-lox site for
insertion into lambda and for site-specific recombination between lox
sites.
Figure 2. Plasmid maps of two fungal expression vectors. Each
plasmid carries a polylinker with 5 or 6 unique restriction sites flanked
on the right by a fungal promoter directing transcription into the
polylinker and flanked on the left by the trpC terminator. Both
plasmids carry a bar selectable marker and a lox-NotI-lox site for
insertion into lambda and for site-specific recombination between lox
sites.