INTRODUCTION
Physical knowledge of the genome increased in parallel with the growth of genetic knowledge. Barbara McClintock showed in 1945 that the seven tiny Neurospora chromosomes can be identified individually using light microscopy (Frontispiece; Fig. 2). She and J. R. Singleton went on to describe chromosome morphology and behavior during meiosis and mitosis in the ascus (1307, 1918) (Fig. 3). Genetic evidence of chromosome rearrangements was confirmed cytologically (907, 1307, 1918). Genetically mapped rearrangements soon enabled linkage groups to be assigned to each of the seven cytologically distinguishable chromosomes (1580). The discovery of insertional and quasiterminal rearrangements made it possible to map genes by duplication coverage [see ref. (1566)]. The complete meiotic karyotype was reconstructed in three dimensions using the synaptonemal complex with its associated recombination nodules (740). Electrophoretic separation of whole-chromosome DNAs provided estimates of the DNA content of individual chromosomes and yielded a value of 43 megabases for the entire haploid genome (1501, 1505) � about three times that of yeast and roughly one-third that of Drosophila or Arabidopsis. Physical mapping (54) and genome sequencing are now in progress.
When the first Neurospora compendium was prepared in 1982, a gene could not be recognized until a phenotypically recognizable variant had been obtained, either as a mutant in the laboratory or as a novel allele from the wild. Molecular genetics has changed all that. Genes can now be recognized on the basis of nucleotide sequence and mapped using restriction fragment length polymorphisms, all before any phenotype is known. Numerous genes in the present compilation have been identified using the cloned wild-type allele. To determine the phenotype of such a gene, a null allele or a mutant allele must be obtained by gene replacement or by repeat-induced point mutation (RIP). Genes without known mutant phenotypes may soon outnumber those for which the mutant phenotype has been determined.
Accounts of the knowledge obtained using Neurospora, and evaluations of the significance of Neurospora for genetics and molecular biology are given in refs. (459)- (461), (893a), (894), and (1575). The development of Neurospora cytogenetics and chromosome cytology is outlined in refs. (461), (1573), (1575), (1578), and (1580). Reference (16691669) provides an overview of the Neurospora genome, with an emphasis on molecular aspects.
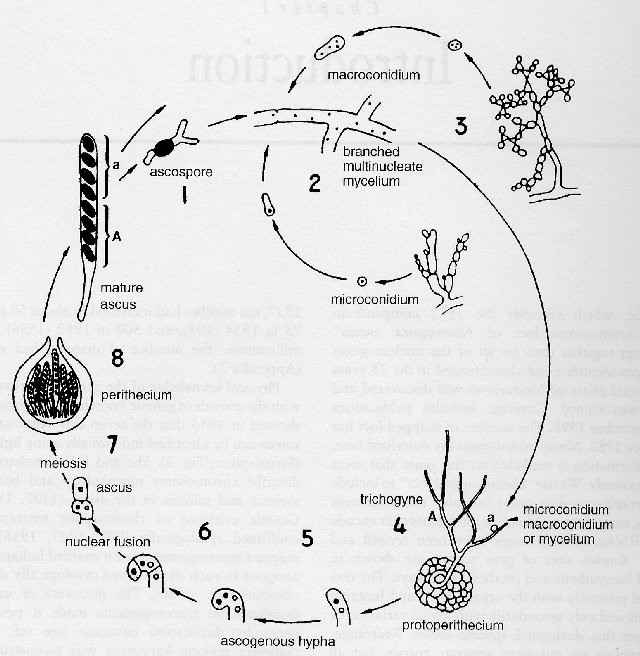
FIGURE 1 The Neurospora crassa life cycle. Stages 1-4, from ascospore germination to fertilization, constitute the vegetative phase, and stages 4-8, from fertilization through ascospore maturation, constitute the sexual phase. Nuclear fusion and meiotic prophase occur 4-5 days after fertilization at 25�C. Ascospores are shot from the perithecium from the ninth day. (1) The black ascospores, approximately 17 x 26 mm, can be isolated manually from an agar surface without the use of micromanipulation apparatus. Ascospore dormancy is broken by heat or chemicals. A germinating ascospore is shown, with hyphae growing from both ends. (2) The mycelium consists of branched, threadlike hyphae made up of multinucleate cells separated by perforate cross-walls through which nuclei and cytoplasm can pass readily. Linear growth of hyphae can exceed 5 mm/h at 37�C. (3) Vegetative spores are of two developmentally distinct types: powdery orange conidia 6-8 mm in diameter, mostly multinucleate, and smaller microconidia, which are uninucleate. (4) Protoperithecia are formed by coiling of hyphae around ascogonial cells. Specialized hyphae, the trichogynes, are attracted to cells of the opposite mating type, from which they pick up nuclei and transport them to the ascogonium within the protoperithecium. Fertilizing nuclei may originate from macroconidia, microconidia, or mycelium. Usually only one fertilizing nuclear type contributes to the contents of a perithecium, but exceptions can occur and are revealed when the fertilizing parent is heterokaryotic (1431) . (5) Upon fertilization, the haploid A and a nuclei do not fuse but proliferate in heterokaryotic ascogenous hyphae. (6) The final conjugate division before ascus formation occurs in a binucleate hook-shaped structure, the crozier. Nuclei of opposite mating type fuse, and the diploid zygote nucleus immediately enters meiosis. The fusion nucleus is the only diploid nucleus in the life cycle. Until ascospore walls are laid down following a postmeiotic mitosis, the ascus remains a single, undivided cell within which the two parental genomes are present in a 1:1 ratio in a common cytoplasm. (7) Details of meiosis and ascus development are diagrammed in Fig. 3 and documented photographically in ref. (1676 ). Asci within a perithecium do not develop synchronously. Numerous asci are initiated successively from the same ascogenous hypha. Mature eight-spored asci measure about 20 x 200 mm. (8) The perithecial wall is maternal in origin. Each mature perithecium, 400-600 mm in diameter, develops a beak that terminates in an osteole through which ascospores are forcibly shot in groups of eight, each comprising the contents of an ascus. Adapted from ref. (645).
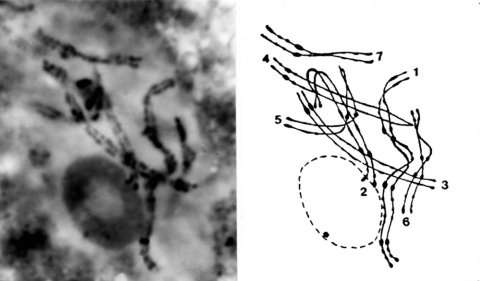
FIGURE 2 The Neurospora crassa chromosome complement in a pachytene nucleus. Following the convention used by McClintock, the chromosomes are numbered in order of their measured length at pachytene. Chromosome 5 has been shifted to the right in the drawing to avoid a confusing overlap with parts of 2 and 4. The overlap in the actual nucleus was not a problem because the arms were in different focal planes. The chromosome 1 bivalent is homozygous for inversion H4250. Orcein staining, X 4000. Photograph by E. G. Barry from ref. (1580).
Stocks and Clones
The preferred source of genetically defined cultures is the Fungal Genetics Stock Center (FGSC),
Department of Microbiology, University of Kansas Medical Center, Kansas City, KS 66160-7420. The FGSC specializes
in Neurospora stocks for research. Over 7500 Neurospora stocks are maintained in the main collection, including
key mutant alleles at most of the known gene loci. Every effort is made to ensure authenticity and, where possible,
to provide stocks with defined genetic backgrounds and free of modifiers and extraneous genes. In addition,
FGSC maintains strains from specialized collections, including >4000 isolates from wild populations. Stock lists are
available on line, and a catalog of the main collection is published biennially as a supplement to
Fungal Genetics Newsletter. Cultures are listed under the following headings: Single Mutants, Multiple Mutants,
Nonchromosomal Mutants, Wild-Type and Wild-Collected Strains (these represent all the known species), Chromosome Rearrangements, and
Reference Strains, Testers, and Stocks for Special Purposes. The last category includes linkage testers, standard
wild types and mating-type testers, diagnostic testers for determining species, heterokaryon-incompatibility testers, strains for
mutant enrichment and replication, progeny from crosses for RFLP mapping, strains for obtaining protoplasts or spheroplasts,
strains for disruption of essential genes, strains for directed transformation, strains for assaying aneuploidy, strains
that produce only microconidia, special strains for teaching, strains for photobiology and circadian rhythms, Spore-killer
testers, transport mutants, and strains for studying mutations in the ad-3 region.
FGSC also has more than 15,000 clones, including EST banks, individual cloned genes, and a growing number of libraries,
including seven genomic libraries, nine cDNA libraries, and three two-hybrid libraries.
The FGSC web site provides links to other culture collections that include Neurospora in their holdings.
The American Type Culture Collection maintains approximately 200 Neurospora stocks, most of which are redundant
with those in FGSC.
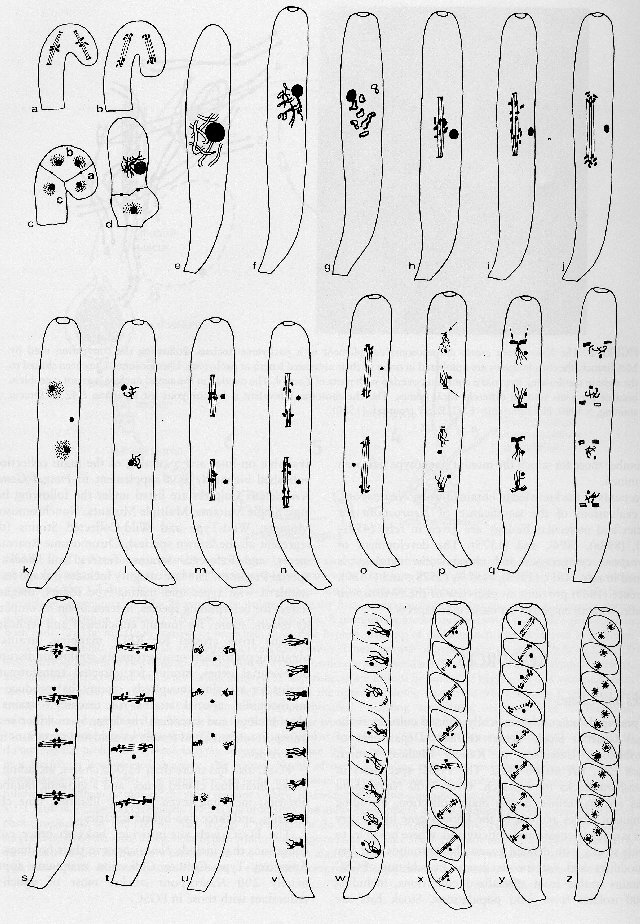
FIGURE 3 Ascus development, meiosis, and ascospore formation in Neurospora crassa. Successive drawings are not to the same scale. (a)-(d) are enlarged several times relative to later stages. Mature ascospores are at least 2-fold larger than ascospores that have just been formed. For clarity, fewer than seven chromosomes are shown in most nuclei. The nuclear envelope (not shown) remains intact during each division. (a) Conjugate metaphase in the crozier. (b) Conjugate telophase. (c) Three-celled crozier. The binucleate penultimate cell (b) grows into an ascus as in (d). The apical cell (a) fuses with the stalk cell (c). (d) Young ascus after nuclear fusion. (e) Pachytene ascus with extended (paired) chromosomes and enlarged nucleolus (see Fig. 2). (f) Diffuse diplotene. The ascus pore forms at this stage. (g) Diakinesis. (h) Metaphase I. (i) Anaphase I. Chromosome separation is asynchronous. (j) Telophase I. (k) Interphase I. (l) Prophase II. (m) Metaphase II. (n) Anaphase II. (o) Telophase II. (p) Early interphase II. Spindle plaques have just formed (arrows). (q) Late interphase II with enlarged double spindle plaques (arrows). (r) Prophase III. Spindle plaques have separated. (s) Metaphase III. Spindles orient across the ascus. (t) Anaphase III. (u) Telophase III. (v) Interphase III. All eight nuclei have lined up in single file. (w) Ascospore delimitation. (x) Metaphase IV (one ascospore is at anaphase IV). (y) Telophase IV. (z) Binucleate ascospores. From ref. (1676), with permission from Urban & Fischer Verlag.
Sources of information
The Internet (1) http://www.fgsc.net (Fungal Genetics Stock Center, Kansas City, KS) This site provides: The FGSC Catalog, including Neurospora wild types, mutants, strains from natural populations, and strains for testing, reference, and other special purposes. The Neurospora holdings are predominantly N. crassa, but other Neurospora species are included, as well as some representatives of related genera.
Lists of available cloned genes and gene libraries.
Fungal Genetics Newsletter (full text for recent years). Includes new acquisitions by FGSC, recent publications, and current genetic maps.
Cumulative RFLP map data.
A bibliography of publications on Neurospora from 1843 to the present. This is a nearly complete listing that combines the 1965 bibliography of Bachmann and Strickland (81) with annual bibliographies from all of the Neurospora Newsletters and Fungal Genetics Newsletters issued in subsequent years.
The complete 1982 Neurospora compendium from Microbiological Reviews 46: 426-570.
Abstracts of Neurospora Conferences and Fungal Genetics Conferences.
Forms for on-line ordering from FGSC and forms for depositing strains and plasmids.
Other features, including descriptions of methods and provisions for discussion and the posting of news. Users of this compendium are encouraged to send corrections and additions to FGSC for posting at their web site.
Information in the FGSC web site is available in formats both for searching and for browsing. Links are provided to other useful sites. A European mirror of the FGSC site will be found at the U.K. Human Genome Mapping Project Resource Centre (HGMP) http://www.hgmp.mrc.ac.uk/research../index.html. (note added in proof: offline in 2002)
(2) http://www.unm.edu/~ngp/ (University of New Mexico Neurospora Genome Project) This site provides an annotated proteome (549a) with searchable databases of EST sequences for genes expressed at different stages in the Neurospora life cycle. (See Appendix 5 for the January 2000 version.) The project also provides and maintains an RFLP mapping program to facilitate placement of new genes using cumulative data for 18 ordered tetrads from a highly polymorphic cross (1452). The RFLP tabulation can be found at the FGSC web site. (See Appendix 3 for the January 2000 version.)
(3) http://www.genome.ou.edu/ (University of Oklahoma Advanced Center for Genome Technology) EST sequences are listed for Neurospora genes expressed at dawn and dusk in the circadian cycle. (4) http://websvr.mips.biochem.mpg.de/ (Munich Information Centre for Protein Sequences) (a) http://websvr.mips.biochem.mpg.de/cgi-bin/proj/medgen/homecrassa Includes information linking Neurospora genes to sequences, with literature citations. (b) http://websvr.mips.biochem.mpg.de/proj/medgen/mitop/ Consolidates information on both nuclear- and mitochondrial-encoded genes and their proteins. Files are given for N. crassa and four other species. Interrelated sections include "Gene Catalogues", "Protein Catalogues," "Homologies," and "Pathways and Metabolism." (c) http://www.mips.biochem.mpg.de/proj/neurospora/ Neurospora genomic DNA sequences for linkage groups II and V. (5) http://fungus.genetics.uga.edu (Fungal Genome Resource, University of Georgia) This site provides data on the physical mapping of the Neurospora genome. Published Sources of Information The most generally useful publication will be Rowland Davis's comprehensive book, "Neurospora" (461). Other useful publications are the book, "Fungal Genetics," by Fincham, Day, and Radford (645) (basic background information); and reviews by Davis and de Serres (462) (basic methods and media) and Perkins and Barry (1580) (cytogenetics). Also useful are the Fungal Genetics Newsletter and the 1982 compendium from Microbiological Reviews (1596). (These are also available at the FGSC web site.) The 1965 bibliography of Bachmann and Strickland (1580) is notable for its meticulously prepared subject index.
Meetings
A Neurospora Conference is held at 2-year intervals, alternating with the more inclusive biennial Fungal Genetics Conference. An elected Neurospora Policy Committee is responsible for coordination and planning of the Neurospora Conference. Committee members are identified in the current Fungal Genetics Newsletter.